Temporal and spatial overlap between monarch larvae and corn pollen
Karen S. Oberhauser*†, Michelle D. Prysby*, Heather R. Mattila‡, Diane E. Stanley-Horn‡, Mark K. Sears‡, Galen Dively§, Eric Olson§, John M. Pleasants¶, Wai-Ki F. Lami , and Richard L. Hellmich**
*University of Minnesota, Department of Ecology, Evolution and Behavior, 1987 Upper Buford Circle, St. Paul, MN 55108; ‡University of Guelph, Department
of Environmental Biology, Guelph, ON, Canada N1G 2W1; §University of Maryland, College Park, MD 20740; ¶Department of Zoology and Genetics, Iowa
State University, Ames, IA 50011; i
Southwest Purdue Agricultural Program, Vincennes, IN 47591; and **United States Department of Agriculture,
Agricultural Research Service (DOA) Corn Insects and Crop Genetics Research Unit, Iowa State University, Ames, IA 50011
To assess the likelihood that monarch larvae will be exposed to
Bacillus thuringiensis (Bt) pollen, we studied milkweed and monarch densities in habitats which comprise much of the land available to breeding monarchs, e.g., cornfields, cornfield edges, other
agricultural fields, and nonagricultural areas, in four regions of the
monarch breeding range. We found that monarchs use milkweed
in cornfields throughout their breeding season, and that per plant
densities are as high or higher in agricultural habitats as in
nonagricultural habitats. As a result of the prevalence of agricultural land, most of the monarchs produced in the upper Midwest
are likely to originate in cornfields or other agricultural habitats.
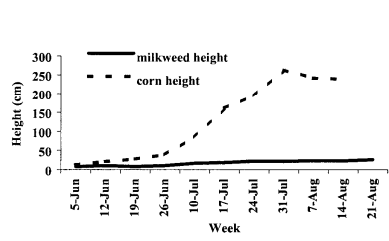
There was a greater temporal overlap between susceptible monarchs and corn anthesis in the northern than the southern part of
the summer breeding range, because of earlier pollen shed in the
south. The importance of agricultural habitats to monarch production suggests that, regardless of the impact of genetically modified
crops, agricultural practices such as weed control and foliar insecticide use could have large impacts on monarch populations.
Transgenic Bacillus thuringiensis (Bt) corn (Zea mays L.) plants
were designed to control the European corn borer [Ostrinia
nubilalis (Hu¨bner)] and other Lepidoptera feeding on corn tissue.
The Bt toxins produced by transgenic corn are lepidopteranspecific and kill only insects that ingest the plant tissue. Because of
this specificity, the impact of Bt corn on nontarget organisms was
assumed to be negligible (1–3). However, most commercial Bt
hybrids express the endotoxin in their pollen to varying degrees and
thus may impose risks to nontarget Lepidoptera that consume
pollen deposited on their host plants (4, 5).
Many aspects of monarch butterfly (Danaus plexippus L.) biology may make them particularly susceptible to impacts of corn pollen exposure. Monarch larvae are present in the late summer, when a portion of the corn acreage is pollinating. A recent stable isotope study (6) suggested the Corn Belt is the origin of most monarchs that migrate to Mexico and form the nucleus of the following year’s population. Malcolm et al. (7) estimated that 92% of the monarchs overwintering in Mexico had fed as larvae on Asclepias syriaca (common milkweed), a common weed in agricultural habitats (8–10).
To determine the potential impact of Bt corn on monarch populations, we need to know the proportion of the monarch population that overlaps temporally and spatially with corn pollen during the larva stage. To estimate this quantity, we measured relative monarch densities in different habitats and the temporal overlap between monarch larvae and corn anthesis. In addition, we compared survivorship of larvae in different habitats to determine whether oviposition patterns reflect monarch production from each habitat.